
Histoire de changer de sujet, un peu de philosophie de la biologie ! Je vais essayer d’aborder une question passionnante et omniprésente en philosophie des sciences : celle de la relation entre les sciences spéciales (chimie, biologie, psychologie…) et la physique fondamentale. Est-ce que les sciences spéciales sont réductibles à la physique ou totalement autonomes ? Si on réduit tout à la physique (« réductionnisme »), est-ce qu’on ne nie pas l’intérêt et le caractère scientifique des sciences spéciales ?

Bien évidemment, il s’agit d’un débat tellement large que je serais bien incapable d’aborder tous ses aspects. Comme pour mes deux billets sur le réalisme scientifique, je vais me contenter de présenter un sujet un peu annexe mais qui permettra d’expliquer des arguments importants autour du réductionnisme. Cela concerne la notion de « mécanisme » : une explication en termes de mécanismes est-elle « réductive » ?
Les mécanismes sont absolument partout en biologie : le mécanisme de réplication de l’ADN, le mécanisme de traduction de l’ARN en protéines, le mécanisme de la réponse immunitaire, les mécanismes cérébraux qui permettent le langage, la mémorisation, la vision, etc… Dans l’article « Thinking about Mechanisms » datant de 2000, les philosophes des sciences Machamer, Darden et Craver (« MDC ») vont jusqu’à dire que « si on ne réfléchit pas en terme de mécanismes, on ne peut pas comprendre la neurobiologie et la biologie moléculaire » (p24). Cette affirmation semble d’autant plus justifiée que la recherche et découverte de mécanismes est très proche de la manière dont travaillent les biologistes et neuroscientifiques.
Face à l’utilisation croissante des mécanismes au sein de la pratique scientifique en biologie, les philosophes ont récemment cherché (à partir des années 2000) à clarifier la signification de ce concept et particulièrement ses implications par rapport à d’autres débats classiques en philosophie des sciences, notamment celui du réductionnisme.
Pour commencer, ce serait pas mal de définir ce qu’est un mécanisme, non ? Et pourquoi est-il si pertinent de penser de la sorte pour comprendre la biologie ?
Le cadre mécaniste défini par Machamer, Darden et Craver (« MDC »)
Toujours dans l’article fondateur « Thinking about mechanisms », « MDC » définissent le mécanisme comme un ensemble « d’entités et d’activités organisées de telle manière à produire des changements réguliers », c’est-à-dire la réalisation d’un phénomène donné à partir de conditions initiales typiques.
Par exemple, quand un agent infectieux s’introduit dans notre organisme, des cellules immunitaires détectent l’intrus et recrutent d’autres cellules sur le site de l’infection via la libération de médiateurs chimiques, entraînant alors une réaction inflammatoire (rougeur, douleur, chaleur…).
Le phénomène de réaction inflammatoire est donc expliqué par le mécanisme consistant en l’activité et l’intéraction des cellules immunitaires (« entités ») présentes dans les tissus.
Ainsi, les activités produisent le changement (ordre temporel, durée, rythme) et font intervenir des entités qui ont certaines propriétés (structure, géométrie, localisation). Le but est de décrire chaque étape intermédiaire du mécanisme de telle sorte qu’il y ait une continuité entre les conditions initiales et le phénomène final. Il ne doit pas y avoir de « vide » dans l’explication : les activités permettent de faire le lien entre les étapes successives du mécanisme.
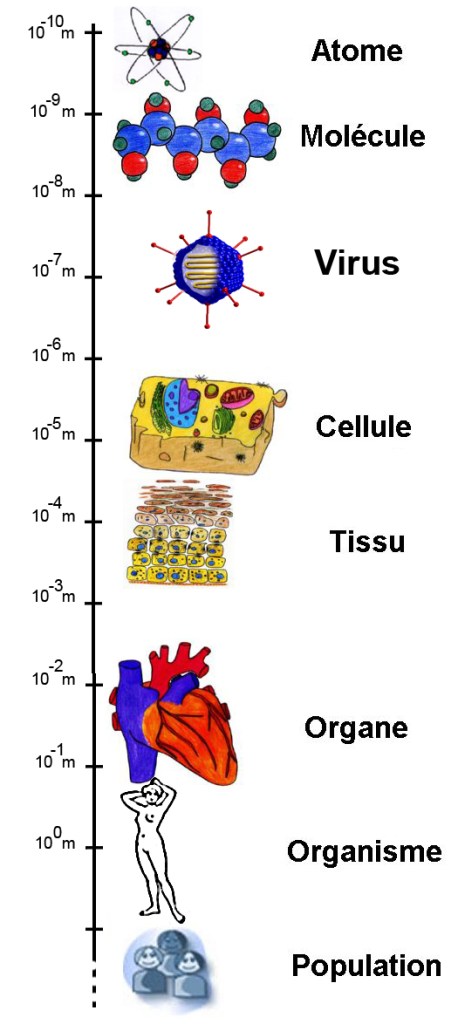
Une autre propriété importante des mécanismes est qu’ils sont « imbriqués » les uns dans les autres. Les explications en biologie se font à plein d’échelles différentes, tant au niveau moléculaire et cellulaire qu’au niveau des comportements des organismes, voire des populations et des espèces. Par exemple, on peut passer des échelles les plus petites à des échelles supérieures : certains mécanismes sont à l’origine de la formation des liaisons chimiques, les liaisons chimiques font partie des mécanismes de réplication de l’ADN, l’ADN est un composant du mécanisme de synthèse des protéines, les protéines interviennent dans tout un tas de processus au sein des cellules, les cellules permettent aux organes d’exercer leur fonction correctement, le cerveau est un organe qui peut nous servir à apprendre la philosophie des sciences, etc, etc.
On voit alors apparaître l’avantage du « cadre mécaniste » selon Craver : les descriptions en termes de mécanismes ont pour ambition d’unifier la biologie, avec l’intégration des explications à toutes les échelles. Le but est également d’éviter de « réduire » les disciplines qui s’intéressent aux couches « supérieures » (échelle des organismes, des comportements des individus…) à celles qui étudient les couches « inférieures » (biologie moléculaire, cellulaire…). Pour préciser un peu, il y a l’ambition d’être « métaphysiquement neutre » : on ne se prononce pas sur la « nature » des différentes couches, l’important est seulement de prendre en compte chacune de leur spécificité pour rendre intelligible une grande variété de phénomènes.
Penser en termes de mécanismes semble alors tout à fait pertinent : les explications sont intuitives, cohérentes avec la pratique scientifique et métaphysiquement neutres, tout en évitant un réductionnisme qui ne reconnaîtrait pas l’intérêt des différents niveaux d’abstraction utilisés en biologie.
Malheureusement… ce n’est pas si simple ! Il y a pas mal d’arguments qui remettent en question ce cadre mécaniste. Lesquels ? Eh bien, c’est là qu’il va falloir approfondir les liens entre mécanisme et réductionnisme !
La démarche mécaniste est-elle vraiment métaphysiquement neutre ?
Comment se positionne le mécanisme par rapport au réductionnisme ? On va voir qu’en fait, la démarche mécaniste ressemble fortement à un certain type de réductionnisme qui a des implications métaphysiques pas du tout neutres.
Pour que ce soit plus facile de le constater, il convient avant de définir un peu plus formellement ce qu’est un mécanisme. En biologie, contrairement à la physique fondamentale, on classe les entités et les phénomènes selon leur fonction caractéristique (ou « rôle causal ») et non selon leur composition physique. Par exemple, le rôle causal du coeur est de pomper le sang à travers l’organisme ; la fonction de certains gènes est de coder pour la couleur des yeux ; etc…
Considérons maintenant un phénomène p que l’on cherche à expliquer en neurosciences : par exemple, la mémorisation spatiale. Cette capacité p est caractérisée par un « rôle causal » ψ : elle nous permet de nous repérer dans des lieux qu’on a déjà visités. Un mécanisme sous-jacent m qui explique p est alors un ensemble de constituants c(1), c(2), …, c(n), caractérisés chacun par leurs activités/« rôles causaux » respectifs, et organisés de telle manière à ce qu’ils reproduisent ensemble le rôle fonctionnel ψ de notre phénomène de départ p. J’ai réalisé ce schéma qui j’espère clarifie la situation :
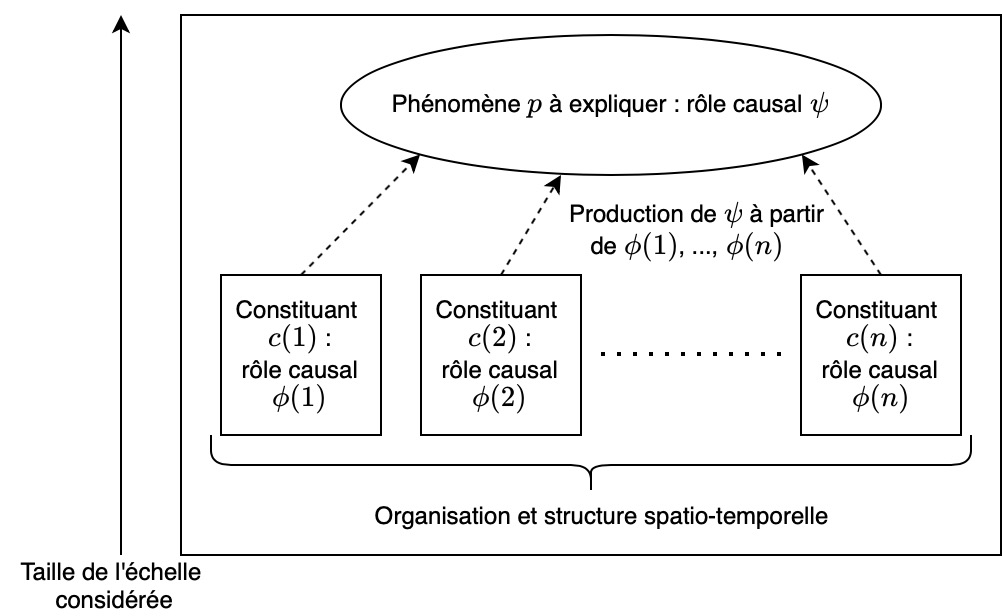
Dans le cas de la mémorisation spatiale, cela reviendrait à étudier la structure du cerveau en identifiant les zones qui contribuent à ce que nous soyons capables de nous repérer dans un lieu connu. Il s’avère par exemple que l’activité de l’hippocampe a un rôle crucial dans notre capacité à représenter mentalement notre environnement, ce qui suggère qu’il serait un constituant c(1) du mécanisme expliquant la mémorisation spatiale.
Se pose alors la question de comment déterminer les constituants d’un mécanisme. Comment distinguer une partie du cerveau qui a un rôle dans la mémorisation spatiale, d’une autre partie qui n’en a pas ? C’est ici que rentre en jeu un critère très important proposé par Craver : c’est le critère de « manipulabilité mutuelle ». Ce nom peut paraître barbare mais l’idée est assez simple. Une entité e est un constituant du mécanisme m qui explique un phénomène global p si et seulement si :
– un changement du comportement de e entraîne une modification du comportement de p, ou dit autrement, un changement du rôle causal de e engendre une modification du rôle causal de p. C’est l’approche « bottom-up » (« du bas vers le haut »). Par exemple, une modification de l’activité de l’hippocampe entraîne une perturbation de la mémorisation spatiale.
– inversement, un changement du comportement global de p entraîne une modification du comportement de e : un changement du rôle causal de p engendre une modification du rôle causal de e. C’est cette fois-ci l’approche « top-down » (« du haut vers le bas »). Par exemple, une modification de notre capacité à nous repérer dans des lieux connus entraîne une perturbation de l’activité de l’hippocampe.
Les comportements des constituants du mécanisme et du phénomène global peuvent donc être manipulés mutuellement, d’où l’expression « manipulabilité mutuelle ».
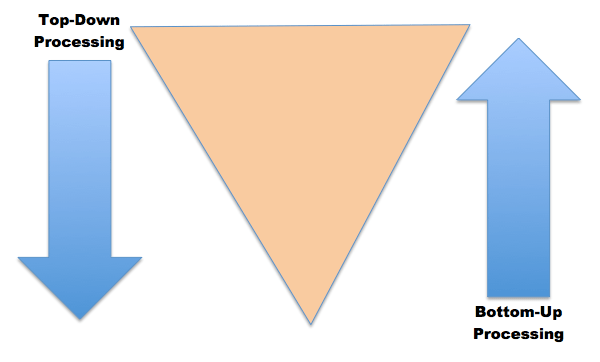
Ce critère proposé par Craver paraît pertinent puisqu’il colle parfaitement à la pratique des scientifiques qui réalisent des expériences « bottom-up » et « top-down » pour identifier les différentes parties du cerveau responsables de l’existence de phénomènes comme la mémorisation spatiale.
Une fois que les constituants c(1), c(2), …, c(n) du mécanisme m ont été trouvés, on peut continuer le raisonnement en expliquant par d’autres mécanismes sous-jacents les activités des constituants de m. L’activité de l’hippocampe peut donc elle-même constituer un phénomène p’, situé à une couche inférieure par rapport à p, et caractérisé par un rôle causal ψ’. Il s’agit alors de trouver un autre mécanisme m’ en identifiant des composants du cerveau d(1), d(2), …., d(N) à un niveau encore inférieur, de telle manière à ce que leurs activités et intéractions reproduisent le rôle fonctionnel ψ’ du phénomène p’. Dans le cas de l’hippocampe, sans rentrer dans les détails, ce sont certaines activités au niveau des synapses qui expliquent en partie la réalisation de la fonction ψ’ du phénomène p’. Et on peut continuer ainsi avec des mécanismes à une couche encore inférieure qui expliquent les activités au niveau des synapses, etc, etc, jusqu’à ce qu’on arrive à l’échelle la plus petite : celle de la physique fondamentale. Comme on a vu précédemment, on obtient finalement des mécanismes imbriqués les uns dans les autres, qui rendent compte des phénomènes à différentes échelles.
Et voilà, on a maintenant toutes les bases nécessaires pour parler du coeur du sujet : le lien entre mécanismes et réductionnisme ! Comme ce billet est déjà assez long et qu’il reste beaucoup de choses à voir, je vais garder ça pour la prochaine fois :). Je devrais normalement publier la suite dans quelques jours. En précisant la relation entre mécanismes et phénomène, on va constater que la démarche mécaniste se rapproche énormément d’un certain type de réductionnisme, le « réductionnisme fonctionnel », et n’échappe donc pas à tout un tas d’implications ontologiques qui sont très problématiques pour la cohérence-même du concept de mécanisme !

Sources principales :
– Machamer, Darden & Craver : Thinking about Mechanisms (2000)
– Patrice Soom : Mechanisms, determination and the metaphysics of neuroscience (2012)
– Stanford Encyclopedia of Philosophy : Mechanisms in Science
(Image ci-contre : le magnifique Nanga Parbat, 8125m)

Ping : Mécanismes et réductionnisme en biologie (2/2) | Science Peak
Très interessant!!
J’aimeJ’aime